《达尔文的危险思想》丹尼特(1995)
第三部分 心灵、意义、数学与道德
第14章 意义的演化
1 追寻真正的意义
约翰·杜威(Dewey, 1910, p. 34):仅仅从运动物质的再分配的角度来说明宇宙的任何做法都不完整,不管它达到了多么真实的程度,因为它忽略了一个基本事实,即运动物质及其再分配的特性在于以累积的方式达到目的——影响我们所知道的价值世界。否认这一点,你就会否认演化论;承认这一点,你就会在目的这个术语唯一客观的,就是唯一可理解的意义上承认它。我并不是说,除了这个机制之外还有其他的理想原因或因素介入。我只是坚持认为,应该注意到这个机制的特征——它就是这样产生并维持各式各样的善的。
我们的意义就像制造品状态的意义一样依赖于功能,因而一样是派生的,一样有潜在的不确定性——有些哲学家无法容忍这种观念,因为它没能让意义具有正规的原因作用(causal role)。我们在前文的一个具体问题中见到过这种观点,它担心心灵仅仅是效果,而不是起始的原因。如果意义是由支持一定功能作用的选择性力量所确定的,那么在某种含义上,意义似乎就不过是在回溯中归结出来的:某个事物的意义并不是它所拥有的一个内在属性,自诞生时起就能对世界有所影响;这个事物的意义顶多是一种回溯性加冕,只有对后续效果加以分析才能确保它的存在。
4 通往未来的安全通道
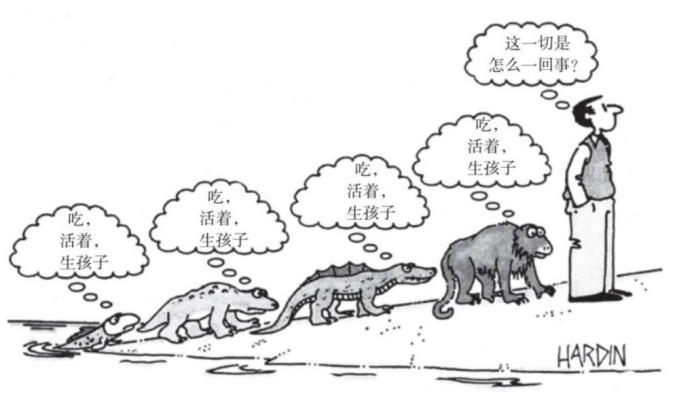
这个机器人仍将只展现出派生的意向性,因为它只是一个制造品,是为了服务你的利益而创造出来的。把这种立场称作“主顾中心主义”。如果你还想坚持这种主顾中心主义,那就应该准备好得出进一步的结论:你自己从来没有享有过任何具有原初意向性的状态,因为你只是一台生存机器,你的设计目的是保存你的基因,直到它们能够复制自己。我们的意向性终归来自我们自私的基因的意向性。它们才是无意的意义赋予者(Unmeant Meaner),而不是我们!
已知有一种足够精致的制造品——某种跟上面这些想象中的机器人同属一类的东西——能够展现出真实的意向性,因为它充分实现了对周边环境的功能性融入,也具有高超的自我保护和自我控制能力。它像你一样,多亏了一个以创造生存机器为目标的规划才会存在,但它也像你一样,已经有了一定的自主性,已经成为一个自我控制、自我决定的场所,这不靠任何奇迹,而只靠面对自己“一生”中的种种难题,并或多或少地解决这些世界摆在它面前的、事关存亡的难题。更为简单的生存机器——比如说设施/植物——永远无法达到你的机器人凭借其复杂性所达到的那种重新定义自我的高度;单单把这些更简单的生存机器看作承载着昏迷居民的生存机器,就可以毫无遗漏地解释它们的所有行为模式。
这样一个机器人会在它的世界中发现意义,会在同他者的交流中开掘这种意义,要承认这种意义就像你所享有的意义一样真实。你的自私的基因就可以被看作你意向性的初始来源——因此也是你所能沉思或想出的每一种意义的初始来源——尽管此后你可以超越你的基因,可以利用你的经验,特别是你所吸收的文化,在你的基因所奠定的基础上建立一个几乎完全独立的(或“超越性的”)意义场所。
一方面,我作为一个人,认为我自己是意义的来源,是判定什么东西重要、为什么重要的仲裁者;而另一方面,我同时也是智人这个物种的一员,是几十亿年的非奇迹研发过程的产物,我们所享有的特征没有一个不是以这样那样的方式从这组过程中产生的。我们的意向性并不会因为它们是数百万年来无心灵的算法研发的后果,而非来自高处的赠予,就不再那么真实。
